Если классифицировать движения организма с точки зрения их биологической значимости для него, то ясно, что на первом плане по значимости окажутся акты, решающие ту или иную возникшую перед особью двигательную задачу. Отсрочивая пока анализ этого понятия, заметим, что значимые задачи, разрешаемые двигательной акцией, как правило, возникают из внешнего окружающего организм мира. Сказанное сразу устраняет из круга значимых акций как все «холостые» движения, не связанные с преодолеванием внешних сил, так и значительную часть мгновенных, однофазных движений типа отдергивания лапы и т.п. Уже отсюда видно, что лабораторная физиология, за малыми исключениями оставлявшая за порогом рабочей комнаты все движения, кроме болевых, оборонительных, самое большее — чесательных рефлексов1, тем самым обедняла свои познавательные ресурсы не только количественно, но и качественно и, как мы сейчас увидим, отнюдь не только в отношении узко двигательной проблематики.
Прежде всего, если относительно «холостых» движений (показывание, проведение линии по воздуху и т.п.) требуются некоторые сведения из механики и биомеханики, чтобы усмотреть для них неотвратимую необходимость кольцевых сенсорных регуляций, то в отношении двигательных актов, сопряженных с преодолением внешних сил, эта необходимость понятна с первого слова. Состоит ли решаемая двигательная задача в локомоции (особенно чем-либо осложненной: бежать по неровному месту, вспрыгивать на возвышение, плыть при волнах и многое другое), в борьбе с другим животным, в рабочем процессе, выполняемом человеком, — всегда предпосылкой для решения является преодоление сил из категории неподвластных, а следовательно, непредусмотримых и не могущих быть преодоленными никаким стереотипом движения, управляемым только изнутри. Неосторожное отметание из поля зрения этих процессов активного взаимодействия с неподвластным окружением (видимо, самоограничение одними «атомами движений» выглядело вполне оправданным для механицистов-атомистов прошлого века, считавших, что целое есть всегда сумма своих частей и ничего более) повело прежде всего к тому, что принцип сенсорной обратной связи, который именно на двигательных объектах мог быть легко усмотрен и обоснован уже 100 лет назад, оставался в тени до недавнего времени.
Долгие годы в физиологии непреоборимо держался в качестве ведущего и универсального принцип разомкнутой рефлекторной дуги. Нельзя исключить возможности того, что действительно в таких элементарных процессах, как рефлекс слюноотделения, или в таких отрывистых и вообще второстепенных по биологическому значению, как рефлекс болевого отдергивания и т.п., дуга не замыкается в рефлекторное кольцо, характерное для схемы управляемого процесса, либо из-за кратковременности акта, либо вследствие его крайней элементарности. Но возможно и вероятно также, что в силу тех же причин краткости и элементарности имеющаяся и здесь циклическая структура ускользала до сих пор от внимания и регистрации (для слюноотделительного процесса это уже почти несомненно). Так или иначе, но представляется очень правдоподобным, что рефлекс по схеме дуги есть лишь рудимент или очень частный случай физиологического реагирования2.
Остается сказать еще об одном ущербе, понесенном физиологией от подмены реальных двигательных актов, разрешающих возникшую объективную задачу, обломками движений почти артефактного характера. Этот последний ущерб, до сего времени не подчеркивавшийся в достаточной степени, очень сильно обеднил наши познания в области рецепторной физиологии и при этом содержал в себе корни важных методологических ошибок.
Нельзя не заметить, что в роли приемника пусковых сигналов, включающих в действие ту или иную рефлекторную дугу, — единственной роли, изучавшейся физиологами классического направления, рецепторные системы, по крайней мере у высокоорганизованных животных и у человека, функционируют существенно и качественно иначе, нежели в роли следящих и корригирующих приборов при выполнении двигательного акта. Это различие можно уяснить, если, став снова на точку зрения биологической значимости, направить внимание на те качества, которые в том и другом случае должны были отсеиваться путем естественного отбора.
Для сигнально-пусковой функции рецептору существенно иметь высокую чувствительность, т.е. максимально низкие пороги как по абсолютной силе сигнала, так и по различению между сигналами. На первый план по биологической значимости здесь выступают телерецепторы обоняния, слуха (также ультраслуха) и зрения в различных ранговых порядках у разных видов животных. Для вычленения далее значимых сигналов из хаотического фона «помех» нужна и необходимо вырабатывается совершенно аналитическая или анализаторная способность реципирующих аппаратов центральной нервной системы (вполне естественно, что И.П.Павлов, в столь большой степени углубивший наши знания по сигнально-пусковой функции рецепторов, присвоил им название анализаторов, только в самые последние годы его жизни дополненное приставкой «синтез»).
Наконец, для этой же сигнально-пусковой роли важнейшим механизмом (который предугадывался уже И.М. Сеченовым и был впоследствии отчетливо экспериментально выявлен исследователями, отправлявшимися от практических задач военного наблюдения) является совокупность процессов активного систематизированного поиска (scanning), или «просматривания», своего диапазона каждым из телерецепторов. Это процессы целиком активные, использующие эффекторику в полной аналогии с тем, как последняя эксплуатирует афферентацию в управлении движениями, но, замечу сразу, не имеющие ничего общего с процессами привлечения организованных двигательных актов к целостному активному восприятию объектов внешнего мира, о чем будет речь дальше.
Когда же двигательный смысловой акт уже «запущен в ход» тем или иным сенсорным сигналом, требования, предъявляемые биологической целесообразностью и приведшие к сформированию в филогенезе механизмов кольцевого сенсорного корригирования, оказываются существенно иными. Что бы ни представляли собой возникшая двигательная задача и тот внешний объект, на который она направлена, для правильной, полезной для особи реализации этой задачи необходимо максимально полное и объективное восприятие как этого объекта, так и каждой очередной фазы и детали собственного действия, направленного к решению данной задачи.
Первая из названных черт рецепторики в этой ее роли — полнота, или синтетичность, обеспечивается хорошо изученными как психо-, так и нейрофизиологами сенсорными синтезами (или сенсорными полями). К их числу относятся, например, схема своего тела, пространственно-двигательное поле, синтезы предметного или «качественного» (топологического) пространства и др. Роль этих «полей» в управлении двигательными актами автор (1947) пытался подробно обрисовать в книге о построении движений. Здесь достаточно будет только напомнить: 1) что в этой функциональной области синтетичность работы рецепторных приборов фигурирует уже не декларативно (как было выше), а как реально прослеженный на движениях в их норме и патологии основной факт и 2) что в каждом из таких сенсорных синтезов, обеспечивающих процессуальное управление двигательными актами, структурная схема объединения между собой деятельности разных проприо-, танго- и телерецепторов имеет свои специфические, качественно и количественно различные свойства. При этом слияние элементарных информаций, притекающих к центральным синтезирующим аппаратам от периферических рецепторов, настолько глубоко и прочно, что обычно почти недоступно расчленению в самонаблюдении. И в описываемой функции принимают участие все или почти все виды рецепторов (может быть, только за исключением вкусового), но уже в существенно иных ранговых порядках. На первом плане оказывается здесь обширная система проприорецепторов в узком смысле. Далее она обрастает соучастием всей танго- и телерецепторики, организовавшейся на основе всего предшествующего практического опыта для выполнения роли «функциональной проприоцепторики». О других, еще только намечающихся чертах чисто физиологического своеобразия работы рецепторов в обсуждаемом круге функций — параметрах адаптации, порогах «по сличению», периодичности функционирования и др. — будет сказано во второй части очерка.
Вторая из названных выше определяющих черт рецепторики как участника кольцевого координационного процесса — объективность — имеет настолько важное принципиальное значение, что на ней необходимо остановиться более подробно.
В той сигнальной (пусковой или тормозной) роли, которая одна только и могла быть замечена при анализе рефлексов по схеме незамкнутой дуги и которая повела к обозначению всего комплекса органов восприятия в центральной нервной системе термином «сигнальная система», от афферентной функции вовсе не требуется доставления объективно верных информаций. Рефлекторная система будет работать правильно, если за каждым эффекторным ответом будет закреплен свой неизменный и безошибочно распознаваемый пусковой сигнал — код. Содержание этого кода, или шифра, может быть совершенно условным, нимало не создавая этим помех к функционированию системы, если только соблюдены два названных сейчас условия. То, что подобный индифферентизм центральной нервной системы к смысловому содержанию сигнала не является каким-то странным, чисто биологическим феноменом, а заложен в самом существе сигнально-пусковой функции, лучше всего доказывается тем, что такими же условными кодированными сигналами безукоризненно осуществляются все необходимые включения и переключения в телеуправляемых автоматах. Можно построить два совершенно одинаковых автомата (самолет-снаряд, мотокатер и т.п.) с одинаковыми моторами, рулями, схемами их радиореле и т.д. и, не внося никаких конструктивных различий, сделать при этом так, чтобы на радиокоды А, Б, В, Г первый отвечал реакциями 1, 2, 3, 4, а второй — реакциями 4,2,1,3 или как угодно иначе.
Cовершенно иными чертами характеризуется работа рецепторной системы при несении ею контрольно-координационных функций по ходу решаемой двигательной задачи. Здесь степень объективной верности информации является решающей предпосылкой для успеха или неуспеха совершаемого действия. На всем протяжении филогенеза животных организмов естественный отбор неумолимо обусловливал отсев тех особей, у которых рецепторы, обслуживавшие их двигательную активность, работали, как кривое зеркало. В ходе онтогенеза каждое столкновение отдельной особи с окружающим миром, ставящее перед собой требующую решения двигательную задачу, содействует, иногда очень дорогой ценой, выработке в ее нервной системе все более верного и точного объективного отражения внешнего мира как в восприятии и осмыслении побуждающей к действию ситуации, так и в проектировке и контроле над реализацией действия, адекватного этой ситуации. Каждое смысловое двигательное отправление, с одной стороны, необходимо требует не условного, кодового, а объективного, количественно и качественно верного отображения окружающего мира в мозгу. С другой стороны, оно само является активным орудием правильного познания этого окружающего мира. Успех или неуспех решения каждой активно пережитой двигательной задачи ведет к прогрессирующей шлифовке и перекрестной выверке показаний упоминавшихся выше сенсорных синтезов и их составляющих3, а также к познанию через действие, проверке через практику, которая является краеугольным камнем всей диалектико-материалистической теории познания, а в разбираемом здесь случае служит своего рода биологическим контекстом к ленинской теории отражения4.
Проведенное на предшествующих страницах сопоставление двух родов функционирования воспринимающих систем организма, пока еще столь неравных по давности их выявления наукой и по степени изученности, позволит осветить несколько по-новому и некоторые черты механизма действия «классических» сигнальных процессов включения или дифференцировочного торможения рефлекторных реакций.
Еще задолго до того, как телемеханика подтвердила существенную принципиальную условность5 пусковых или переключательных кодов, этот же факт был установлен на биологическом материале знаменитым открытием И.П. Павлова. Факт, что любой раздражитель из числа доступных восприятию может быть с одинаковой легкостью превращен в пусковой сигнал для того или другого органического (безусловного) рефлекса, оказался в дальнейшем чрезвычайно универсальным.
Как показали последующие работы павловской школы (А.Д. Сперанский), во всем комплексе физиологических функций, вплоть до самых, казалось бы, недосягаемых глубинных процессов гормонального или клеточно-метаболического характера, нет ни одного отправления, которое не могло бы быть подсоединено (в принципе одним и тем же методом) к любому пусковому раздражителю. Этот замечательный индифферентизм нервных систем к содержанию и качеству пусковых сигналов был отмечен И.П. Павловым уже в самом начальном периоде изучения открытого им круга явлений. Об этом свидетельствует и наименование, данное раздражителям, вновь искусственно прививаемым к стволам старых прирожденных рефлексов, — условные раздражители. Название, предложенное В.М. Бехтеревым, — сочетательные раздражители и рефлексы — менее глубоко в отношении внутреннего смысла явлений, но зато вплотную подводит к схеме их механизмов, которая к нашему времени уяснилась уже вполне отчетливо.
Для превращения любого надпорогового агента в условный пусковой раздражитель того или другого органического рефлекса требуется всегда обеспечение двух условий: 1) главного — встречи или сочетания в пределах обычно небольшого интервала времени этого агента с реализацией данного рефлекса и 2) побочного — некоторого числа повторений такого сочетания. Первое из этих условий прямо относит разбираемый феномен к циклу ассоциаций по смежности, как раз характеризующихся безразличием к смысловому содержанию ассоциируемых представлений или рецепций. Интересно отметить, что для преобразования индифферентного раздражителя в условнопусковой существенно совмещение его с эффекторной, а не с афферентной частью безусловного рефлекса, которая мобилизуется в типовом эксперименте только как средство заставить сработать эффекторную полудугу. Это доказывается, например, фактом осуществимости так называемых условных рефлексов второго порядка, когда индифферентный раздражитель приобретает пусковые свойства для данного рефлекса, несмотря на то что эффекторная часть последнего запускается в действие не безусловным, а ранее привитым к рефлексу условным же раздражителем первого порядка.
Другое доказательство сказанного можно усмотреть в том, что в методах, применяемых при дрессировке, поощрительное подкрепление «безусловным» афферентным импульсом подкормки животного производится после правильного выполнения им требуемого действия по соответствующей условной команде и не является при этом безусловным пусковым раздражителем дрессируемого действия. Эта недооценивавшаяся раньше деталь заслуживает внимания в настоящем контексте потому, что образование ассоциативной связи в мозгу между условным афферентным процессом и эффекторной частью рефлекса, как нам кажется, можно осмыслить, только если эффекторная реализация рефлекса отражается (опять-таки по кольцевой обратной связи) назад в центральную нервную систему и может уже сочетаться ассоциативно с афферентным же процессом условного раздражения. Это могло бы послужить еще одним подтверждением того, что возвратно-афферентационные акты как непосредственные соучастники процесса и в классических рефлексах — «дугах» — не отсутствуют, а лишь пока ускользают от наблюдения.
Второе из условий образования условной связи, названное выше побочным, а именно надобность некоторого числа повторных сочетаний, было бы трудно объяснить сейчас иначе, как необходимостью для подопытной особи выделить прививаемую новую рецепцию из всего хаоса бомбардирующих ее извне воздействий. Число повторений должно оказаться достаточным для того, чтобы определилась неслучайность совмещения во времени интеро- или проприоцепции реализующего рефлекса именно с данным элементом всей совокупности экстерорецепций. В этом смысле — в отношении необходимого и достаточного числа повторений — раздражитель, индифферентный по своему смысловому содержанию, может оказаться относительно труднее и длительнее вычленяемым как могущий не привлечь к себе интереса и внимания («ориентировочной реакции») особи. Старую наивно-материалистическую концепцию о постепенных «проторениях» путей или синаптических барьеров в центральной нервной системе можно уже считать сданной в архивы науки6.
В нескольких словах заслуживает упоминания факт, который и в свете новых приобретений регуляционной физиологии продолжает оставаться неясным. Структура почти всех изучавшихся условных сочетаний такова, что к органической, безусловной эффекторной полудуге прививается новый условный афферентный пусковой сигнал. Разнообразие как безусловных эффекторных процессов, так и афферентных «позывных», которые могут быть привязаны к первым, совершенно безгранично; но неизвестно почти ни одного случая, который обнаружил бы обратную структуру условной связи: прицепление нового условного эффекторного окончания к безусловной афферентной полудуге. До некоторой степени случаи такого обратного типа наблюдались в давнишних опытах М.Е. Ерофеевой (1912), но сам И.П. Павлов7, цитировавший их в своих «Лекциях о работе больших полушарий», сопровождает их описание рядом ограничений и оговорок. Как бы ни был объяснен в дальнейшем такой «структурный парадокс», ясно, что инертная неизменяемость именно эффективной полудуги реально осуществимых условно-двигательных рефлексов чрезвычайно затрудняет использование их структурного механизма для обучения незнакомым движениям, образования и усовершенствования двигательных навыков и сноровок и т.п.
Рассмотрение вопроса о сигнальных кодах и их сочетательной роли в аспекте регуляционной физиологии способно, как нам кажется, по-новому осветить вопрос о так называемой второй сигнальной системе. Из всего проанализированного выше ясно, что при ничем не ограниченном разнообразии возможных условносигнальных кодов в их число могут входить и речевые фонемы, вовсе не образуя в этой своей роли какого-либо особого класса и нуждаясь, как и все раздражения, пригодные для роли сигналов, только в воспринимаемости и распознаваемости. Никто не приписывал собаке, медведю, морскому льву, кошке обладание второй сигнальной системой или архитектоническими полями, гомологичными полю Вернике человека. Между тем любое из этих животных (даже не сплошь «высших» млекопитающих) дрессируется на словесные сигналы с такой же легкостью, с какой образуются у них условные замыкания и дифференцировки на другие виды раздражений. Эти фонематические сигнальные коды, ничем существенным не выделяющиеся из других кодов, могли генетически у первобытного человека явиться зародышами фонем — приказов, своего рода рудиментарными повелительными наклонениями, из которых впоследствии эволюционировали речевые формы глагола8.
C другой стороны, назывательные элементы речи, из которых у человека сформулировалась категория имен, никогда не несли и не могли, разумеется, нести какой-либо сигнальной функции в определенном выше смысле. Поэтому трактовка «второй сигнальной системы» как системы словесного отображения предметов (вообще первичных рецепций внешних объектов, образующих по этой концепции в совокупности «первую сигнальную систему»), что полностью проявляется и в составе словника, применяемого в экспериментах по так называемой речедвигательной методике, представляется результатом глубоко ошибочного смешения двух резко различных физиологических функций и речевых категорий. Слова как сигналы не образуют никакой особой системы и в роли пусковых фонем полностью доступны многим животным, еще чрезвычайно далеким от функции речи. Слова и речь как отражение внешнего мира в его статике (имена) и динамике действий и взаимодействий с субъектом (глаголы, суждения) действительно образуют систему, доступную и свойственную только человеку. Но обозначать речь, достигшую этой ступени значения и развития, как сигнальную систему, — значит подменять ее одним из самых несущественных и рудиментарных ее проявлений9.
Идея второй сигнальной системы, несомненно, явилась одним из следствий упоминавшегося выше методологического ущерба, понесенного физиологией из-за признания ею одной только сигнально-пусковой роли рецепторики и недооценки ее важнейших биологических и социальных функций: познавания через действие и регуляции активного воздействия на окружающий мир. Знак равенства, ставившийся между понятиями рецепции и сигнала, вынуждал относить к категории сигналов и перципируемое слово. Между тем нельзя было пройти мимо огромного качественного своеобразия речи как специфически присущего виду Homo sapiens символического отображения воспринимаемого мира и себя самого в нем. Упоминавшаяся уже выше терпимость к атомизму легко позволила зато пройти мимо структурности речи (делающей ее не скоплением слов, а орудием мышления) и трактовать ее как сумму слов — сигналов преимущественно конкретно-предметного содержания.
Отечественная физиология сумела избегнуть другой, гораздо более важной гносеологической ошибки, в которую легко впадали многие мыслители западного мира и которая также целиком проистекает из одностороннего понимания рецепторной функции: от несомненного факта примиримости безукоризненного функционирования рефлекторных приборов с полной условностью вызывающих их сенсорных кодов очень легко соскользнуть на путь признания символичности всяких вообще рецепций, условности картины мира в мозгу и психике, непознаваемости объективной реальности и прочих идеалистических концепций, давно ниспровергнутых подлинной наукой.
* * *
Перейдем к уточненному анализу механизмов двигательной координации у высших организмов, имея в виду две задачи: 1) извлечь из этого анализа максимум доступных на сегодняшний день указаний на общие закономерности механизмов управления и 2) попытаться выявить, в чем состоит то своеобразие моторики высших животных, в особенности человека, которое резко качественно отграничивает ее действия и ресурсы от всего того, чего мы можем ожидать от автоматной техники сегодняшнего и, вероятно, даже завтрашнего дня. Мне неизбежно придется касаться многих пунктов, в свое время уже подробно проанализированных (Бернштейн, 1946, 1947). Во избежание неуместных повторений я в настоящем очерке буду излагать их как можно более кратко, лишь для соблюдения непрерывности логической линии изложения. Здесь будет более правильным постараться дополнить и углубить затрагивавшиеся там вопросы, преимущественно касающиеся основных, принципиальных механизмов координационного управления, попутно исправляя ошибки, выяснившиеся к настоящему времени.
Первое резкое биомеханическое отличие двигательного аппарата человека и высших животных от любого из искусственных самодействующих устройств, неоднократно подчеркивавшееся, состоит в огромном, выражающемся трехзначными числами количестве доступных ему степеней свободы как кинематических, зависящих от многозвенности его свободно сочлененных кинематических цепей, так и эластических, обусловленных упругостью движущих тяг — мышц и отсутствием в силу этого однозначных отношений между мерой активности мышц, ее напряжением, длиной и скоростью ее изменения. Для уяснения того, как осложняет управление движением каждая лишняя степень свободы, ограничусь здесь двумя примерами.
Cудно на поверхности моря имеет три степени свободы (если пренебречь движениями качки), но практически достаточным является управление одной-единственной степенью — направлением, или курсом, так как на морских просторах, если судно, отклоненное чем-нибудь от курса, восстановит прежнее направление, то ему нет необходимости возвращаться на старую трассу и вполне достаточно продолжать путь параллельно ей, в паре кабельтовых в ту или другую сторону. Эта задача успешно решается компасным автопилотом. Но представим себе автомобиль, который должен ехать по шоссе ограниченной ширины, автоматически выполняя все встречаемые кривизны и повороты. Здесь управлению практически подлежат всего две степени свободы его подвижности.
Анализ показывает, что независимо от способа получения машинной информации о ходе шоссе (относительно, например, его средней линии), будут ли они восприниматься фото-, электро-, механорецепторами и т.д., блок-схема такого автомата рулевого управления, способного вести автомобиль по изгибам шоссе, удерживая его близ средней линии, должна содержать: 1) рецептор расстояния от линии с его знаком, 2) рецептор угла между осью машины и средней линией с его знаком, 3) рецептор фактической кривизны пути, 4) суммирующий смеситель и 5) систему регуляторов для гашения паразитных «болтаний» машины в ту и другую сторону от курса. Столько осложнений вносится в проблему автоматизации всего одною лишней степенью свободы. Насколько мне известно, автомат подобного рода еще никогда не был создан.
Полезно отметить, что огромная трудность его осуществления отнюдь не в сигнализации или устройстве рецепторов названных типов: то и другое имеется и сейчас на вооружении автоматики. Вся трудность состоит в организации центральной перешифровки информаций, получаемых на входе от фотоэлементов или магнитных реле, в качество, силу и последовательность импульсов, управляющих сервомоторами рулевого аппарата.
Второй пример для сопоставления приведу из области нормальной координации движений человека при нормальной работе всех его афферентных органов и лишь в условиях необычной двигательной задачи. Прицепите спереди к пряжке своего пояса верхний конец лыжной палки, на конце, несущем колесико, прикрепите груз в 1-2 кг, а к колесику справа и слева привяжите по резиновой трубке достаточной длины для того, чтобы можно было взять концы каждой в правую и левую руку. Направив палку острием вперед, станьте перед вертикальной доской, на которой крупно начерчен круг, квадрат или иная простая фигура, и постарайтесь, манипулируя только потягиванием за резиновые трубки, обвести острием палки начерченную фигуру. Палка изображает здесь одно звено конечности с двумя кинематическими степенями свободы, трубки -аналоги всех мышц-антагонистов, привносящих в систему еще две упругие степени свободы. Этот опыт (очень демонстративный при его показе в аудитории) убедит каждого, испробовавшего его в роли исследуемого, какая нелегкая и малопослушная для координирования вещь — всего четыре степени свободы, когда к услугам человека, даже находящегося во всеоружии всех своих рецепторов, не имеется моторного опыта, приобретаемого по отношению к костно-мышечному двигательному аппарату с самых первых недель жизни.
Определение координации, данное мной в упоминавшихся работах, кажется мне и сейчас наиболее строгим и точным. Координация движений есть преодоление избыточных степеней свободы движущегося органа, иными словами — превращение его в управляемую систему. Короче, координация есть организация управляемости двигательного аппарата. В основном определении с намерением говорится не о закреплении, протормаживании и т.п. избыточных степеней свободы, а об их преодолении. Как показали экстенсивные работы с детьми<…>, фиксация, устраняющая упомянутый избыток, применяется как наиболее примитивный и невыгодный путь лишь в самом начале осваивания двигательного умения, сменяясь затем более гибкими, целесообразными и экономичными путями преодоления этого избытка через организацию всего процесса. Какую преобладающую роль может играть именно организация регуляционных взаимодействий даже в нехитром случае управления только двумя степенями свободы, мог показать уже наш первый пример с автопилотажем вдоль шоссе.
В своих работах о построении движений и частично в предыдущих очерках этой книги я подробно останавливался на причинах, создающих биодинамическую необходимость организованных по кольцевому принципу механизмов двигательной координации, и на некоторых обнаруженных наблюдением чертах тех физиологических процессов контрольного взаимодействия, которые обеспечивают координационное руководство движением при посредстве сенсорных синтезов разных уровней построения. Там было показано, какое огромное место в ряду непредусмотримых и практически неподвластных сил, требующих непрерывного восприятия и преодоления, занимают наряду с внешними силами реактивные силы, неизбежно возникающие при движениях в многозвенных кинематических цепях органов движения и усложняющиеся в огромной прогрессии с каждым лишним звеном сочленовной цепи и с каждой новой степенью свободы подвижности. Не затрагивая здесь более этой чисто биодинамической стороны проблем, обратимся к вопросу, оставшемуся в тени в названных выше работах, но все более назревающему в ходе современного развития физиологической мысли. Если двигательная координация есть система механизмов, обеспечивающая управляемость двигательного аппарата и позволяющая утилизировать с уверенностью всю его богатую и сложнейшую подвижность, то что можно сказать к настоящему времени о путях и механизмах самого управления двигательными актами? В каких отношениях могут уловимые для нас в настоящее время закономерности этого управления оказаться полезными для интересов прикладной кибернетики (бионики) и какие из сторон или свойств этих закономерностей отсеются как наиболее специфические для нервных систем высших животных и человека и поэтому наиболее способные осветить ту пропасть, которая пока еще (и, видимо, надолго) качественно разделяет достижения автоматики от реализующейся в двигательных актах жизнедеятельности высокоразвитых организмов?
Предварительно нужно вкратце остановиться на некоторых вопросах терминологии и попытаться систематизировать известные на сегодняшний день принципиальные схемы саморегулирующихся устройств (в дальнейшем для краткости мы будем обозначать этот термин первыми буквами — СУ) с интересующих нас проблемных позиций.
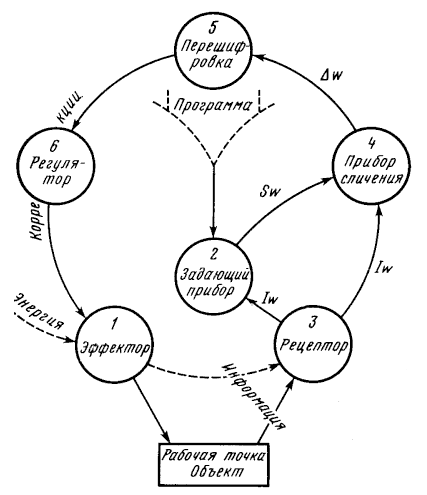
Все системы, саморегулирующиеся по какому-либо параметру, постоянному или переменному, обязаны, как минимум, содержать в своем составе следующие элементы:
1) эффектор (мотор), работа которого подлежит регулированию по данному параметру;
2) задающий элемент, вносящий тем или другим путем в систему требуемое значение регулируемого параметра;
3) рецептор, воспринимающий фактические текущие значения параметра и сигнализирующий о них каким-либо способом в прибор сличения;
4) прибор сличения, воспринимающий расхождение фактического и требуемого значений с его величиной и знаком;
5) устройство, перешифровывающее данные прибора сличения в коррекционные импульсы, подаваемые по обратной связи на регулятор;
6) регулятор, управляющий по данному параметру функционированием эффектора .
Вся система образует, таким образом, замкнутый контур взаимодействий, общая схема которого дана на рис. 1. Между перечисленными элементами нередко бывают включены не имеющие принципиального значения вспомогательные устройства: усилители, реле, сервомоторы и т.п.
В примере, приводимом Wiener по идее его партнера Rosenbluth, координационное управление жестом взятия видимого предмета со стола рассматривается как непрерывная оценка уменьшения того куска пути, какой еще остается пройти кисти руки до намеченного предмета. При всей правомерности обозначения места предмета как Sw, текущего положения кисти — как Iw, а планомерно убывающего расстояния между ними — как переменной Dw = (Iw-Sw) я должен пояснить здесь же, что и выше, и в дальнейшем рассматриваю координационный процесс в микроинтервалах пути и времени, опираясь на данные, собранные за годы работы моей и моих товарищей. Поэтому в рамках настоящего очерка я рассматриваю как переменный Sw весь непрерывный запланированный путь, или процесс движения органа, а как Iw — фактически текущие координаты последнего. В связи с этим Dw в настоящем контексте — это пороговомалые отклонения, корригируемые более или менее исправно по ходу движения. Примером их могут служить отклонения линии, проводимой от руки карандашом или острием планиметра, от начерченной линии, которую требуется обвести. В нашем смысле, следовательно, Dw есть не планомерно убывающая макродистанция, а колеблющаяся, то возникающая, то погашаемая с тем или иным успехом малая величина переменного знака и направления.
Централ